- Bosh sahifa
- Slaydlar
- Fizika
- Oqsil biosintezi. Oqsilning post-translasyonal modifikatsiyasi. Protein sintezining energiya xarajatlari
Oqsil biosintezi. Oqsilning post-translasyonal modifikatsiyasi. Protein sintezining energiya xarajatlari
Yuklangan vaqt:
04.12.2023
Hajmi:
5547.1806640625 KB

![Protein sintezini keng ikki bosqichga bo'lish mumkin - transkripsiya va tarjima. Transkripsiya paytida DNK deb nomlanuvchi oqsilni kodlash gen, deb nomlangan shablon molekulasiga aylantiriladi xabarchi RNK. Ushbu konversiya fermentlar tomonidan amalga oshiriladi RNK polimerazalar, hujayraning yadrosida.[2] Eukaryotlarda bu xabarchi RNK (mRNA) dastlab erta shaklda (pre-mRNK) ishlab chiqariladi va etuk mRNK hosil qilish uchun transkripsiyadan keyingi modifikatsiyaga uchraydi. Etuk mRNK eksport qilinadi yadro orqali yadro teshiklari uchun sitoplazma tarjima sodir bo'lishi uchun hujayraning. Tarjima paytida mRNA o'qiladi ribosomalar ishlatadigan nukleotid ning ketma-ketligini aniqlash uchun mRNK ketma-ketligi aminokislotalar. Ribosomalar hosil bo'lishini katalizlaydi kovalent peptid bog'lari polipeptid zanjirini hosil qilish uchun kodlangan aminokislotalar o'rtasida. Tarjimadan so'ng polipeptid zanjiri buklanib, funktsional oqsil hosil qilishi kerak, masalan, ferment vazifasini o'tashi uchun polipeptid zanjiri funktsional hosil bo'lishi uchun to'g'ri katlanishi kerak. faol sayt. Funktsional uch o'lchovli (3D) shaklni qabul qilish uchun polipeptid zanjiri avval bir qator kichik pastki tuzilmalarni hosil qilishi kerak ikkilamchi tuzilmalar. Keyinchalik ushbu ikkilamchi tuzilmalardagi polipeptid zanjiri buklanib, umumiy 3D hosil qiladi uchinchi darajali tuzilish.](/data/documents/d6e09bae-4245-4dad-8db4-7bab0e14e6f4/page-3.png)
![To'g'ri katlangandan so'ng, oqsil turli xil pishib yetishi mumkin tarjimadan keyingi modifikatsiyalar. Translyatsiyadan keyingi modifikatsiyalar oqsilning ishlash qobiliyatini, hujayra ichida joylashgan joyni (masalan, sitoplazma yoki yadro) va oqsilning qobiliyatini o'zgartirishi mumkin. boshqa oqsillar bilan ta'sir o'tkazish. Protein biosintezi kasallikda asosiy rol o'ynaydi, chunki bu jarayonda o'zgarishlar va xatolar, asosiy sabab DNK mutatsiyalari yoki oqsilning noto'g'ri birikishi, ko'pincha kasallikning asosiy sabablari hisoblanadi. DNK mutatsiyalari keyingi mRNK ketma-ketligini o'zgartiradi va keyinchalik mRNA kodlangan aminokislotalar ketma-ketligini o'zgartiradi. Mutatsiyalar polipeptid zanjirining qisqarishiga olib kelishi mumkin hosil qilish orqali to'xtash ketma-ketligi bu tarjimaning erta tugatilishiga olib keladi. Shu bilan bir qatorda, mRNA ketma-ketligidagi mutatsiya shu holatda kodlangan o'ziga xos aminokislotani o'zgartiradi polipeptid zanjirida Ushbu aminokislotaning o'zgarishi oqsillarning ishlashiga yoki to'g'ri katlanmasına ta'sir qilishi mumkin.[4] Noto'g'ri katlanmış oqsillar hosil bo'lish uchun bir-biriga yopishib qolish xususiyatiga ega bo'lganligi sababli, qoniqarsiz oqsillar ko'pincha kasallikka chalinadi zich protein to'plamlari. Ushbu birikmalar ko'pincha bir qator kasalliklar bilan bog'liq nevrologik, shu jumladan Altsgeymer kasalligi va Parkinson kasalligi.](/data/documents/d6e09bae-4245-4dad-8db4-7bab0e14e6f4/page-4.png)
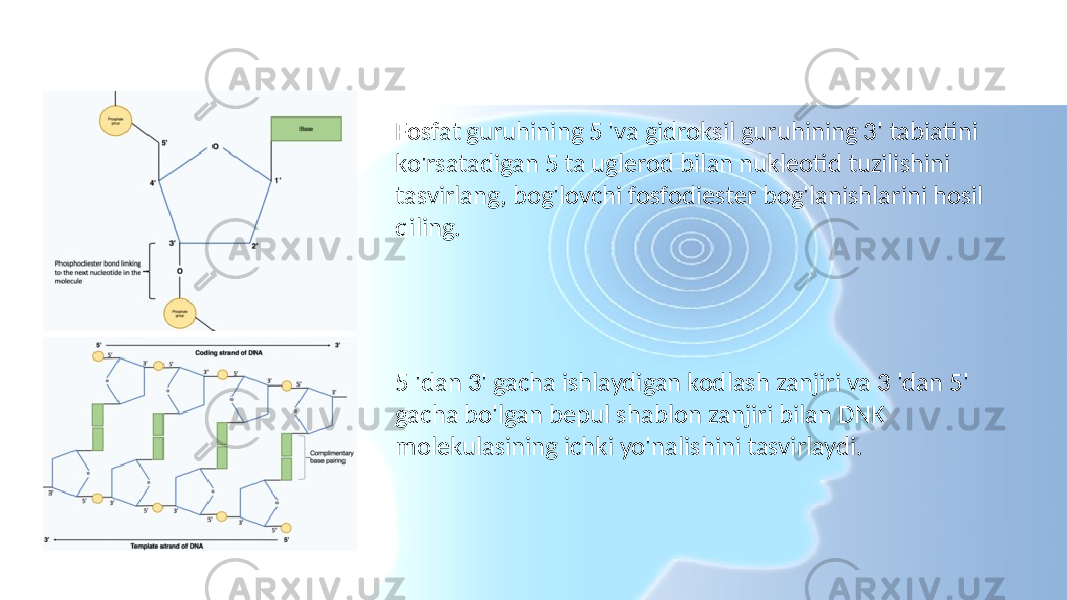

![RNK polimeraza tomonidan DNKning shablon zanjirining mRNKgacha molekulasiga aylanishini tasvirlaydi. Ferment RNK polimeraza ochiq shablon ipiga bog'lanib, gendan 3 'dan 5' gacha o'qiydi. Bir vaqtning o'zida RNK polimeraza 5 m dan 3 'yo'nalishda mRNKgacha bo'lgan bitta zanjirni hosil bo'lishini katalizlab sintez qiladi. fosfodiester aloqalari bir-birini to'ldirishga qodir bo'lgan faol yadro (nukleotidlar) o'rtasida asosiy juftlik shablon ipi bilan. Harakatlanuvchi RNK-polimeraza ortida DNKning ikkita zanjiri qayta qo'shiladi, shuning uchun bir vaqtning o'zida faqat 12 bazaviy juft DNK ta'sir qiladi.[6] RNK polimeraza mRNKgacha molekulasini soniyada 20 nukleotid tezligida hosil qiladi, shu bilan bir soat ichida bir xil gendan minglab mRNKgacha molekulalarni ishlab chiqarish imkoniyatini beradi. Sintezning tez sur'atlariga qaramay, RNK polimeraza fermenti o'zining tuzatish mexanizmini o'z ichiga oladi. Korrektor mexanizmlari RNK-polimeraza eksizion reaktsiyasi orqali o'sib boruvchi mRNK molekulasidan noto'g'ri nukleotidlarni (DNKning shablon zanjiriga qo'shimcha bo'lmagan) olib tashlashga imkon beradi. [1] RNK polimerazalar ma'lum bir DNK ketma-ketligiga yetganda tugaydi transkripsiya, RNK polimeraza ajralib chiqadi va mRNKgacha sintez tugaydi.[6] Sintez qilingan mRNKgacha molekulasi shablon DNK zanjiri bilan to'ldiruvchidir va kodlash DNK zanjiri bilan bir xil nukleotidlar qatoriga ega. Ammo DNK va mRNK molekulalarining nukleotid tarkibida bitta muhim farq mavjud. DNK asoslardan tashkil topgan - guanin, sitozin, adenin va timin (G, C, A va T) - RNK shuningdek to'rt asosdan iborat - guanin, sitozin, adenin va urasil. RNK molekulalarida DNK asos timini o'rnini adenin bilan juftlashtirishga qodir uratsil egallaydi. Shuning uchun mRNKgacha bo'lgan molekulada kodlash DNK zanjirida timin bo'lgan barcha qo'shimcha asoslar uratsil bilan almashtiriladi. [7]](/data/documents/d6e09bae-4245-4dad-8db4-7bab0e14e6f4/page-7.png)
![RNK polimeraza tomonidan DNKning shablon zanjirining mRNKgacha molekulasiga aylanishini tasvirlaydi. Ferment RNK polimeraza ochiq shablon ipiga bog'lanib, gendan 3 'dan 5' gacha o'qiydi. Bir vaqtning o'zida RNK polimeraza 5 m dan 3 'yo'nalishda mRNKgacha bo'lgan bitta zanjirni hosil bo'lishini katalizlab sintez qiladi. fosfodiester aloqalari bir-birini to'ldirishga qodir bo'lgan faol yadro (nukleotidlar) o'rtasida asosiy juftlik shablon ipi bilan. Harakatlanuvchi RNK-polimeraza ortida DNKning ikkita zanjiri qayta qo'shiladi, shuning uchun bir vaqtning o'zida faqat 12 bazaviy juft DNK ta'sir qiladi.[6] RNK polimeraza mRNKgacha molekulasini soniyada 20 nukleotid tezligida hosil qiladi, shu bilan bir soat ichida bir xil gendan minglab mRNKgacha molekulalarni ishlab chiqarish imkoniyatini beradi. Sintezning tez sur'atlariga qaramay, RNK polimeraza fermenti o'zining tuzatish mexanizmini o'z ichiga oladi. Korrektor mexanizmlari RNK- polimeraza eksizion reaktsiyasi orqali o'sib boruvchi mRNK molekulasidan noto'g'ri nukleotidlarni (DNKning shablon zanjiriga qo'shimcha bo'lmagan) olib tashlashga imkon beradi.[1] RNK polimerazalar ma'lum bir DNK ketma-ketligiga yetganda tugaydi transkripsiya, RNK polimeraza ajralib chiqadi va mRNKgacha sintez tugaydi.[6]](/data/documents/d6e09bae-4245-4dad-8db4-7bab0e14e6f4/page-8.png)

![Transkripsiyadan keyingi modifikatsiyaning uchta asosiy bosqichi mavjud: 1) A qo'shilishi 5 'shapka mRNKgacha bo'lgan molekulaning 5 'oxirigacha 2) 3 'qo'shilishi poli (A) quyruq mRNKgacha bo'lgan 3 'uchi molekulasiga qo'shiladi 3) Olib tashlash intronlar orqali RNK qo'shilishi 5 'kepka mRNKgacha bo'lgan molekulaning 5' uchiga qo'shiladi va orqali o'zgartirilgan guanin nukleotididan iborat. metilatsiya. 5 'qopqoqning maqsadi tarjima qilishdan oldin etuk mRNK molekulalarining parchalanishini oldini olishdir, shuningdek qopqoq ribosomaning mRNK bilan bog'lanishiga yordam beradi. [8] va mRNKni hujayradagi boshqa RNKlardan ajratib olishga imkon beradi.[1] Aksincha, 3 'Poly (A) dumi mRNK molekulasining 3' uchiga qo'shiladi va 100-200 adenin asosidan iborat.[8] Ushbu aniq mRNA modifikatsiyalari hujayraning 5 'shapka va 3' dumlari bo'lsa ham mRNA xabarining to'liqligini aniqlashga imkon beradi.[1] Ushbu modifikatsiyalangan mRNKgacha bo'lgan molekula keyinchalik RNK qo'shilish jarayonidan o'tadi. Genlar bir qator intronlardan va exons, intronlar - bu oqsilni kodlamaydigan nukleotidlar ketma-ketligi, ekzonlar - oqsilni to'g'ridan-to'g'ri kodlaydigan nukleotidlar. Intronlar va ekzonlar asosiy DNK ketma-ketligida ham, mRNKgacha bo'lgan molekulada ham mavjud, shuning uchun oqsilni kodlovchi etuk mRNK molekulasini hosil qilish uchun birikish kerak.[6] Splichlash jarayonida oraliq intronlar mRNKdan oldingi molekuladan a deb nomlanuvchi ko'p oqsilli kompleks tomonidan olib tashlanadi. splitseozoma (150 dan ortiq oqsil va RNKdan iborat).[9] Ushbu etuk mRNK molekulasi keyinchalik yadro konvertidagi yadro teshiklari orqali sitoplazma ichiga eksport qilinadi.](/data/documents/d6e09bae-4245-4dad-8db4-7bab0e14e6f4/page-10.png)
